A l’aube du développement de la vie sur Terre, toutes les formes cellulaires étaient représentées par des bactéries. Ils absorbaient les substances organiques dissoutes dans l’océan primordial à travers la surface du corps.
Au fil du temps, certaines bactéries se sont adaptées pour produire des substances organiques à partir de substances inorganiques. Pour ce faire, ils ont utilisé l’énergie du soleil. Le premier système écologique est apparu dans lequel ces organismes étaient producteurs. En conséquence, l’oxygène libéré par ces organismes est apparu dans l’atmosphère terrestre. Avec son aide, vous pouvez obtenir beaucoup plus d'énergie du même aliment et utiliser l'énergie supplémentaire pour compliquer la structure du corps : diviser le corps en parties.
L’une des réalisations importantes de la vie est la séparation du noyau et du cytoplasme. Le noyau contient des informations héréditaires. Une membrane spéciale autour du noyau permettait de se protéger contre les dommages accidentels. Selon les besoins, le cytoplasme reçoit des commandes du noyau qui dirigent la vie et le développement de la cellule.
Les organismes dans lesquels le noyau est séparé du cytoplasme ont formé le superrègne nucléaire (ceux-ci comprennent les plantes, les champignons et les animaux).
Ainsi, la cellule - base de l'organisation des plantes et des animaux - est née et s'est développée au cours de l'évolution biologique.
Même à l'œil nu, ou mieux encore à la loupe, on peut constater que la chair d'une pastèque mûre est constituée de très petits grains, ou grains. Ce sont des cellules – les plus petits « éléments constitutifs » qui constituent le corps de tous les organismes vivants, y compris les plantes.
La vie d'une plante s'effectue grâce à l'activité combinée de ses cellules, créant un tout unique. Avec la multicellularité des parties végétales, il existe une différenciation physiologique de leurs fonctions, une spécialisation de diverses cellules en fonction de leur localisation dans le corps végétal.
Une cellule végétale diffère d’une cellule animale en ce qu’elle possède une membrane dense qui recouvre le contenu interne de tous les côtés. La cellule n'est pas plate (comme on le représente habituellement), elle ressemble très probablement à une très petite bulle remplie de contenu muqueux.
Structure et fonctions d'une cellule végétale
Considérons une cellule comme une unité structurelle et fonctionnelle d'un organisme. L’extérieur de la cellule est recouvert d’une paroi cellulaire dense dans laquelle se trouvent des sections plus minces appelées pores. En dessous se trouve un film très fin – une membrane recouvrant le contenu de la cellule – le cytoplasme. Dans le cytoplasme se trouvent des cavités - des vacuoles remplies de jus cellulaire. Au centre de la cellule ou près de la paroi cellulaire se trouve un corps dense - un noyau avec un nucléole. Le noyau est séparé du cytoplasme par l'enveloppe nucléaire. De petits corps appelés plastes sont répartis dans tout le cytoplasme.
Structure d'une cellule végétale
Structure et fonctions des organites des cellules végétales
| Organoïde | Dessin | Description | Fonction | Particularités |
Paroi cellulaire ou membrane plasmique | Incolore, transparent et très résistant | Fait passer des substances dans et hors de la cellule. | La membrane cellulaire est semi-perméable |
|
Cytoplasme | Substance visqueuse épaisse | Toutes les autres parties de la cellule s'y trouvent | Est en mouvement constant |
|
Noyau (partie importante de la cellule) | Rond ou ovale | Assure le transfert des propriétés héréditaires aux cellules filles lors de la division | Partie centrale de la cellule |
|
De forme sphérique ou irrégulière | Participe à la synthèse des protéines | |||
 | Réservoir séparé du cytoplasme par une membrane. Contient de la sève cellulaire | Les nutriments de rechange et les déchets dont la cellule n’a pas besoin s’accumulent. | Au fur et à mesure que la cellule grandit, les petites vacuoles fusionnent en une seule grande vacuole (centrale). |
|
Plastides | Chloroplastes | Ils utilisent l'énergie lumineuse du soleil et créent des matières organiques à partir d'éléments inorganiques. | La forme des disques délimités du cytoplasme par une double membrane |
|
Chromoplastes | Formé à la suite de l’accumulation de caroténoïdes | Jaune, orange ou marron |
||
 | Leucoplastes | Plastides incolores | ||
Enveloppe nucléaire | Se compose de deux membranes (externe et interne) avec des pores | Sépare le noyau du cytoplasme | Permet les échanges entre le noyau et le cytoplasme |
La partie vivante d'une cellule est un système ordonné et structuré de biopolymères et de structures membranaires internes impliqués dans un ensemble de processus métaboliques et énergétiques qui maintiennent et reproduisent l'ensemble du système dans son ensemble.
Une caractéristique importante est que la cellule ne possède pas de membranes ouvertes aux extrémités libres. Les membranes cellulaires limitent toujours les cavités ou zones, les fermant de tous côtés.
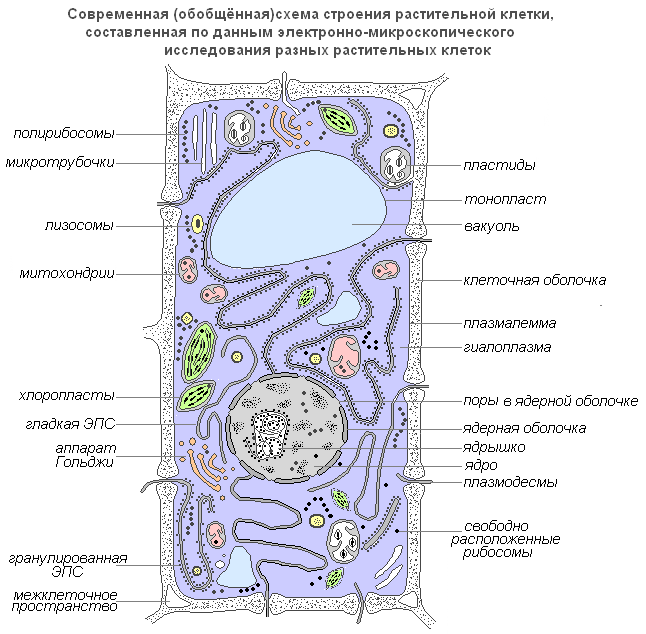
Schéma généralisé moderne d'une cellule végétale
Plasmalemme(membrane cellulaire externe) est un film ultramicroscopique de 7,5 nm d'épaisseur, constitué de protéines, de phospholipides et d'eau. Il s'agit d'un film très élastique, bien mouillé par l'eau et qui rétablit rapidement son intégrité après un dommage. Il a une structure universelle, c'est-à-dire typique de toutes les membranes biologiques. Dans les cellules végétales, à l’extérieur de la membrane cellulaire se trouve une paroi cellulaire solide qui crée un support externe et maintient la forme de la cellule. Il est constitué de fibres (cellulose), un polysaccharide insoluble dans l'eau.
Plasmodesmes les cellules végétales, sont des tubules submicroscopiques qui pénètrent dans les membranes et sont tapissés d'une membrane plasmique, qui passe ainsi d'une cellule à l'autre sans interruption. Avec leur aide, une circulation intercellulaire de solutions contenant des nutriments organiques se produit. Ils transmettent également des biopotentiels et d’autres informations.
Porami appelées ouvertures dans la membrane secondaire, où les cellules ne sont séparées que par la membrane primaire et la lame médiane. Les zones de la membrane primaire et de la plaque médiane séparant les pores adjacents des cellules adjacentes sont appelées membrane poreuse ou film de fermeture du pore. Le film de fermeture du pore est percé de tubules plasmodesmiques, mais aucun trou traversant n'est généralement formé dans les pores. Les pores facilitent le transport de l'eau et des solutés d'une cellule à l'autre. Des pores se forment dans les parois des cellules voisines, généralement les unes en face des autres.
Membrane cellulaire a une coque bien définie et relativement épaisse de nature polysaccharidique. L'enveloppe d'une cellule végétale est le produit de l'activité du cytoplasme. L'appareil de Golgi et le réticulum endoplasmique participent activement à sa formation.

Structure de la membrane cellulaire
La base du cytoplasme est sa matrice, ou hyaloplasme, un système colloïdal complexe, incolore et optiquement transparent, capable de transitions réversibles du sol au gel. Le rôle le plus important de l'hyaloplasme est d'unir toutes les structures cellulaires en un seul système et d'assurer l'interaction entre elles dans les processus du métabolisme cellulaire.
Hyaloplasme(ou matrice cytoplasmique) constitue l'environnement interne de la cellule. Il est constitué d'eau et de divers biopolymères (protéines, acides nucléiques, polysaccharides, lipides), dont l'essentiel est constitué de protéines de spécificité chimique et fonctionnelle variable. Le hyaloplasme contient également des acides aminés, des monosaccharides, des nucléotides et d'autres substances de faible poids moléculaire.
Les biopolymères forment un milieu colloïdal avec l'eau qui, selon les conditions, peut être dense (sous forme de gel) ou plus liquide (sous forme de sol), à la fois dans tout le cytoplasme et dans ses sections individuelles. Dans le hyaloplasme, divers organites et inclusions sont localisés et interagissent entre eux et avec l'environnement hyaloplasmique. De plus, leur localisation est le plus souvent spécifique à certains types de cellules. A travers la membrane bilipide, le hyaloplasme interagit avec l'environnement extracellulaire. Par conséquent, l’hyaloplasme est un environnement dynamique et joue un rôle important dans le fonctionnement des organites individuels et dans la vie des cellules en général.
Formations cytoplasmiques - organites
Les organites (organites) sont des composants structurels du cytoplasme. Ils ont une certaine forme et taille et constituent des structures cytoplasmiques obligatoires de la cellule. S’ils sont absents ou endommagés, la cellule perd généralement sa capacité à continuer d’exister. De nombreux organites sont capables de se diviser et de s’auto-reproduire. Leurs tailles sont si petites qu’elles ne peuvent être observées qu’au microscope électronique.
Cœur
Le noyau est l’organite le plus important et généralement le plus grand de la cellule. Il a été étudié pour la première fois en détail par Robert Brown en 1831. Le noyau assure les fonctions métaboliques et génétiques les plus importantes de la cellule. Sa forme est assez variable : elle peut être sphérique, ovale, lobée ou en forme de lentille.
Le noyau joue un rôle important dans la vie de la cellule. Une cellule dont le noyau a été retiré ne sécrète plus de membrane et cesse de croître et de synthétiser des substances. Les produits de décomposition et de destruction s'y intensifient, ce qui entraîne sa mort rapide. La formation d'un nouveau noyau à partir du cytoplasme ne se produit pas. Les nouveaux noyaux se forment uniquement en divisant ou en écrasant les anciens.
Le contenu interne du noyau est la caryolymphe (suc nucléaire), qui remplit l'espace entre les structures du noyau. Il contient un ou plusieurs nucléoles, ainsi qu'un nombre important de molécules d'ADN liées à des protéines spécifiques - les histones.

Structure de base
Nucléole
Le nucléole, comme le cytoplasme, contient majoritairement de l'ARN et des protéines spécifiques. Sa fonction la plus importante est de former des ribosomes, qui réalisent la synthèse des protéines dans la cellule.
Appareil de Golgi
L'appareil de Golgi est un organite universellement distribué dans tous les types de cellules eucaryotes. Il s'agit d'un système à plusieurs niveaux de sacs membranaires plats qui s'épaississent le long de la périphérie et forment des processus vésiculaires. Elle est le plus souvent située à proximité du noyau.

Appareil de Golgi
L'appareil de Golgi comprend nécessairement un système de petites vésicules (vésicules), qui se détachent de citernes épaissies (disques) et sont situées le long de la périphérie de cette structure. Ces vésicules jouent le rôle de système de transport intracellulaire de granules sectoriels spécifiques et peuvent servir de source de lysosomes cellulaires.
Les fonctions de l'appareil de Golgi consistent également en l'accumulation, la séparation et la libération en dehors de la cellule à l'aide de vésicules de produits de synthèse intracellulaire, de produits de désintégration et de substances toxiques. Les produits de l'activité synthétique de la cellule, ainsi que diverses substances entrant dans la cellule depuis l'environnement par les canaux du réticulum endoplasmique, sont transportés vers l'appareil de Golgi, s'accumulent dans cet organite, puis sous forme de gouttelettes ou de grains pénètrent dans le cytoplasme et sont soit utilisés par la cellule elle-même, soit excrétés à l'extérieur. Dans les cellules végétales, l'appareil de Golgi contient des enzymes pour la synthèse des polysaccharides et le matériau polysaccharidique lui-même, qui est utilisé pour construire la paroi cellulaire. On pense qu’il participe à la formation des vacuoles. L'appareil de Golgi doit son nom au scientifique italien Camillo Golgi, qui l'a découvert pour la première fois en 1897.
Lysosomes
Les lysosomes sont de petites vésicules délimitées par une membrane dont la fonction principale est d'effectuer la digestion intracellulaire. L'utilisation de l'appareil lysosomal se produit lors de la germination d'une graine de plante (hydrolyse des nutriments de réserve).

Structure d'un lysosome
Microtubules
Les microtubules sont des structures membraneuses et supramoléculaires constituées de globules protéiques disposés en spirale ou en rangées droites. Les microtubules remplissent une fonction principalement mécanique (motrice), assurant la mobilité et la contractilité des organites cellulaires. Situés dans le cytoplasme, ils donnent à la cellule une certaine forme et assurent la stabilité de la disposition spatiale des organites. Les microtubules facilitent le mouvement des organites vers des endroits déterminés par les besoins physiologiques de la cellule. Un nombre important de ces structures sont localisées dans le plasmalemme, à proximité de la membrane cellulaire, où elles participent à la formation et à l'orientation des microfibrilles de cellulose des parois cellulaires végétales.

Structure des microtubules
Vacuole
La vacuole est le composant le plus important des cellules végétales. C'est une sorte de cavité (réservoir) dans la masse du cytoplasme, remplie d'une solution aqueuse de sels minéraux, d'acides aminés, d'acides organiques, de pigments, de glucides et séparée du cytoplasme par une membrane vacuolaire - le tonoplaste.
Le cytoplasme remplit toute la cavité interne uniquement dans les cellules végétales les plus jeunes. Au fur et à mesure que la cellule grandit, la disposition spatiale de la masse initialement continue de cytoplasme change de manière significative : de petites vacuoles remplies de sève cellulaire apparaissent et la masse entière devient spongieuse. Avec la poursuite de la croissance cellulaire, les vacuoles individuelles fusionnent, poussant les couches de cytoplasme vers la périphérie, de sorte que la cellule formée contient généralement une grande vacuole et que le cytoplasme avec tous les organites est situé près de la membrane.
Les composés organiques et minéraux solubles dans l'eau des vacuoles déterminent les propriétés osmotiques correspondantes des cellules vivantes. Cette solution d'une certaine concentration est une sorte de pompe osmotique permettant une pénétration contrôlée dans la cellule et la libération de l'eau, des ions et des molécules métabolites.
En combinaison avec la couche cytoplasmique et ses membranes, caractérisées par des propriétés semi-perméables, la vacuole forme un système osmotique efficace. Les indicateurs de cellules végétales vivantes tels que le potentiel osmotique, la force d'aspiration et la pression de turgescence sont déterminés de manière osmotique.

Structure de la vacuole
Plastides
Les plastes sont les plus grands organites cytoplasmiques (après le noyau), inhérents uniquement aux cellules des organismes végétaux. On ne les trouve pas uniquement dans les champignons. Les plastes jouent un rôle important dans le métabolisme. Ils sont séparés du cytoplasme par une double membrane et certains types possèdent un système de membranes internes bien développé et ordonné. Tous les plastes sont de la même origine.
Chloroplastes- les plastes les plus courants et les plus importants sur le plan fonctionnel des organismes photoautotrophes qui effectuent des processus photosynthétiques, conduisant finalement à la formation de substances organiques et à la libération d'oxygène libre. Les chloroplastes des plantes supérieures ont une structure interne complexe.

Structure chloroplastique
Les tailles des chloroplastes dans différentes plantes ne sont pas les mêmes, mais leur diamètre moyen est de 4 à 6 microns. Les chloroplastes sont capables de se déplacer sous l'influence du mouvement du cytoplasme. De plus, sous l'influence de l'éclairage, on observe un mouvement actif des chloroplastes de type amiboïde vers la source lumineuse.
La chlorophylle est la substance principale des chloroplastes. Grâce à la chlorophylle, les plantes vertes sont capables d'utiliser l'énergie lumineuse.
Leucoplastes(plastes incolores) sont des corps cytoplasmiques clairement définis. Leurs tailles sont légèrement inférieures à celles des chloroplastes. Leur forme est également plus uniforme, se rapprochant de la sphérique.

Structure leucoplastique
Trouvé dans les cellules épidermiques, les tubercules et les rhizomes. Lorsqu'ils sont illuminés, ils se transforment très rapidement en chloroplastes avec une modification correspondante de la structure interne. Les leucoplastes contiennent des enzymes à l'aide desquelles l'amidon est synthétisé à partir de l'excès de glucose formé lors de la photosynthèse, dont l'essentiel se dépose dans les tissus ou organes de stockage (tubercules, rhizomes, graines) sous forme de grains d'amidon. Chez certaines plantes, les graisses se déposent dans les leucoplastes. La fonction de réserve des leucoplastes se manifeste parfois par la formation de protéines de réserve sous forme de cristaux ou d'inclusions amorphes.
Chromoplastes dans la plupart des cas, ce sont des dérivés de chloroplastes, parfois de leucoplastes.

Structure chromoplastique
La maturation des cynorrhodons, des poivrons et des tomates s'accompagne de la transformation des chloro- ou leucoplastes des cellules pulpaires en plastes caratinoïdes. Ces derniers contiennent principalement des pigments plastidiques jaunes - des caroténoïdes, qui, à maturité, y sont intensément synthétisés, formant des gouttelettes lipidiques colorées, des globules solides ou des cristaux. Dans ce cas, la chlorophylle est détruite.
Mitochondries
Les mitochondries sont des organites caractéristiques de la plupart des cellules végétales. Ils ont une forme variable de bâtons, de grains et de fils. Découvert en 1894 par R. Altman à l'aide d'un microscope optique, la structure interne a ensuite été étudiée à l'aide d'un microscope électronique.

La structure des mitochondries
Les mitochondries ont une structure à double membrane. La membrane externe est lisse, la membrane interne forme des excroissances de formes diverses - des tubes dans les cellules végétales. L'espace à l'intérieur de la mitochondrie est rempli d'un contenu semi-liquide (matrice), qui comprend des enzymes, des protéines, des lipides, des sels de calcium et de magnésium, des vitamines, ainsi que de l'ARN, de l'ADN et des ribosomes. Le complexe enzymatique des mitochondries accélère le mécanisme complexe et interconnecté des réactions biochimiques qui aboutissent à la formation d'ATP. Ces organites fournissent de l'énergie aux cellules - la conversion de l'énergie des liaisons chimiques des nutriments en liaisons à haute énergie de l'ATP au cours du processus de respiration cellulaire. C'est dans les mitochondries que se produit la dégradation enzymatique des glucides, des acides gras et des acides aminés avec la libération d'énergie et sa conversion ultérieure en énergie ATP. L'énergie accumulée est dépensée en processus de croissance, en nouvelles synthèses, etc. Les mitochondries se multiplient par division et vivent environ 10 jours, après quoi elles sont détruites.
Réticulum endoplasmique
Le réticulum endoplasmique est un réseau de canaux, tubes, vésicules et citernes situés à l'intérieur du cytoplasme. Découvert en 1945 par le scientifique anglais K. Porter, il s'agit d'un système de membranes à structure ultramicroscopique.

Structure du réticulum endoplasmique
L'ensemble du réseau est réuni en un seul tout avec la membrane cellulaire externe de l'enveloppe nucléaire. Il existe des ER lisses et rugueux, qui portent des ribosomes. Sur les membranes du RE lisse se trouvent des systèmes enzymatiques impliqués dans le métabolisme des graisses et des glucides. Ce type de membrane prédomine dans les cellules graines riches en substances de stockage (les protéines, les glucides, les huiles sont attachées à la membrane granulaire du RE, et lors de la synthèse d'une molécule protéique, la chaîne polypeptidique avec les ribosomes est immergée dans le canal ER. Les fonctions du réticulum endoplasmique sont très diverses : transport de substances aussi bien au sein de la cellule qu'entre cellules voisines ; division d'une cellule en sections distinctes dans lesquelles divers processus physiologiques et réactions chimiques se déroulent simultanément.
Ribosomes
Les ribosomes sont des organites cellulaires non membranaires. Chaque ribosome est constitué de deux particules qui ne sont pas de taille identique et peuvent être divisées en deux fragments, qui continuent de conserver la capacité de synthétiser des protéines après s'être combinés en un ribosome entier.

Structure des ribosomes
Les ribosomes sont synthétisés dans le noyau, puis le quittent et se déplacent vers le cytoplasme, où ils sont attachés à la surface externe des membranes du réticulum endoplasmique ou sont localisés librement. Selon le type de protéine synthétisée, les ribosomes peuvent fonctionner seuls ou être combinés en complexes - les polyribosomes.
(nucléaire). Les cellules procaryotes ont une structure plus simple ; apparemment, elles sont apparues plus tôt dans le processus d’évolution. Les cellules eucaryotes sont plus complexes et sont apparues plus tard. Les cellules qui composent le corps humain sont eucaryotes.
Malgré la variété des formes, l'organisation des cellules de tous les organismes vivants est soumise à des principes structurels communs.
Cellule procaryote
Cellule eukaryotique
Structure d'une cellule eucaryote
Complexe superficiel d'une cellule animale
Comprend glycocalice, membranes plasmiques et la couche corticale du cytoplasme située en dessous. La membrane plasmique est également appelée plasmalemme, la membrane externe de la cellule. Il s'agit d'une membrane biologique d'environ 10 nanomètres d'épaisseur. Assure avant tout une fonction délimitatrice par rapport à l'environnement extérieur à la cellule. De plus, il remplit une fonction de transport. La cellule ne gaspille pas d'énergie pour maintenir l'intégrité de sa membrane : les molécules sont maintenues ensemble selon le même principe que les molécules de graisse - il est thermodynamiquement plus avantageux que les parties hydrophobes des molécules soient situées à proximité immédiate. les uns aux autres. Le glycocalice est constitué de molécules d’oligosaccharides, de polysaccharides, de glycoprotéines et de glycolipides « ancrées » dans le plasmalemme. Le glycocalyx remplit des fonctions de récepteur et de marqueur. La membrane plasmique des cellules animales est principalement constituée de phospholipides et de lipoprotéines entrecoupées de molécules protéiques, notamment d'antigènes de surface et de récepteurs. Dans la couche corticale (adjacente à la membrane plasmique) du cytoplasme se trouvent des éléments cytosquelettiques spécifiques - des microfilaments d'actine ordonnés d'une certaine manière. La fonction principale et la plus importante de la couche corticale (cortex) réside dans les réactions pseudopodiales : éjection, fixation et contraction des pseudopodes. Dans ce cas, les microfilaments sont réarrangés, allongés ou raccourcis. La forme de la cellule (par exemple la présence de microvillosités) dépend également de la structure du cytosquelette de la couche corticale.
Structure cytoplasmique
Le composant liquide du cytoplasme est également appelé cytosol. Au microscope optique, il semblait que la cellule était remplie de quelque chose comme du plasma liquide ou du sol, dans lequel le noyau et d'autres organites « flottaient ». En fait, ce n'est pas vrai. L'espace interne d'une cellule eucaryote est strictement ordonné. Le mouvement des organites est coordonné à l'aide de systèmes de transport spécialisés, appelés microtubules, qui servent de « routes » intracellulaires et de protéines spéciales, dynéines et kinésines, qui jouent le rôle de « moteurs ». Les molécules de protéines individuelles ne diffusent pas non plus librement dans tout l’espace intracellulaire, mais sont dirigées vers les compartiments nécessaires à l’aide de signaux spéciaux à leur surface, reconnus par les systèmes de transport de la cellule.
Réticulum endoplasmique
Dans une cellule eucaryote, il existe un système de compartiments membranaires (tubes et citernes) se croisant les uns dans les autres, appelé réticulum endoplasmique (ou réticulum endoplasmique, ER ou EPS). La partie du RE, aux membranes de laquelle les ribosomes sont attachés, est appelée granulaire(ou rugueux) réticulum endoplasmique, la synthèse des protéines se produit sur ses membranes. Les compartiments qui n'ont pas de ribosomes sur leurs parois sont classés comme lisse(ou agranulaire) ER, qui participe à la synthèse des lipides. Les espaces internes du RE lisse et granulaire ne sont pas isolés, mais se croisent et communiquent avec la lumière de l'enveloppe nucléaire.
Appareil de Golgi
Cœur
Cytosquelette
Centrioles
Mitochondries
Comparaison des cellules pro- et eucaryotes
La différence la plus importante entre les eucaryotes et les procaryotes a longtemps été considérée comme la présence d'un noyau formé et d'organites membranaires. Cependant, dans les années 1970-1980. il est devenu clair que ce n'était qu'une conséquence de différences plus profondes dans l'organisation du cytosquelette. Pendant un certain temps, on a cru que le cytosquelette n'était caractéristique que des eucaryotes, mais au milieu des années 1990. des protéines homologues aux principales protéines du cytosquelette des eucaryotes ont également été découvertes chez les bactéries.
C'est la présence d'un cytosquelette spécifiquement structuré qui permet aux eucaryotes de créer un système d'organites mobiles de la membrane interne. De plus, le cytosquelette permet l'endo- et l'exocytose (on suppose que c'est grâce à l'endocytose que les symbiotes intracellulaires, dont les mitochondries et les plastes, sont apparus dans les cellules eucaryotes). Une autre fonction importante du cytosquelette eucaryote est d'assurer la division du noyau (mitose et méiose) et du corps (cytotomie) de la cellule eucaryote (la division des cellules procaryotes s'organise plus simplement). Les différences dans la structure du cytosquelette expliquent également d'autres différences entre les pro- et les eucaryotes - par exemple, la constance et la simplicité des formes des cellules procaryotes et la diversité significative de forme et la capacité de la modifier dans les cellules eucaryotes, ainsi que la taille relativement importante de cette dernière. Ainsi, la taille des cellules procaryotes est en moyenne de 0,5 à 5 microns, celle des cellules eucaryotes est en moyenne de 10 à 50 microns. De plus, ce n'est que parmi les eucaryotes qu'il existe des cellules véritablement géantes, comme les œufs massifs de requins ou d'autruches (dans un œuf d'oiseau, le jaune entier est un énorme œuf), des neurones de grands mammifères dont les processus, renforcés par le cytosquelette , peut atteindre des dizaines de centimètres de longueur.
Anaplasie
La destruction de la structure cellulaire (par exemple dans les tumeurs malignes) est appelée anaplasie.
Histoire de la découverte cellulaire
La première personne à voir des cellules fut le scientifique anglais Robert Hooke (connu grâce à la loi de Hooke). Au cours de l'année, essayant de comprendre pourquoi le liège flotte si bien, Hooke a commencé à examiner de fines sections de liège à l'aide d'un microscope qu'il avait amélioré. Il découvrit que le liège était divisé en de nombreuses petites cellules, qui lui rappelaient les cellules des monastères, et il appela ces cellules cellules (en anglais cell signifie « cellule, cellule, cage »). La même année, le maître néerlandais Anton van Leeuwenhoek (-) utilise pour la première fois un microscope pour voir des « animaux » – des organismes vivants en mouvement – dans une goutte d'eau. Ainsi, dès le début du XVIIIe siècle, les scientifiques savaient que sous un fort grossissement, les plantes avaient une structure cellulaire et ils ont vu certains organismes qui furent plus tard appelés unicellulaires. Cependant, la théorie cellulaire de la structure des organismes ne s'est formée qu'au milieu du XIXe siècle, après l'apparition de microscopes plus puissants et le développement de méthodes de fixation et de coloration des cellules. L'un de ses fondateurs était Rudolf Virchow, mais ses idées contenaient un certain nombre d'erreurs : par exemple, il supposait que les cellules étaient faiblement connectées les unes aux autres et que chacune existait « par elle-même ». Ce n'est que plus tard qu'il a été possible de prouver l'intégrité du système cellulaire.
voir également
- Comparaison de la structure cellulaire des bactéries, des plantes et des animaux
Liens
- Molecular Biology Of The Cell, 4e édition, 2002 - manuel de biologie moléculaire en anglais
- Cytologie et génétique (0564-3783) publie des articles en russe, ukrainien et anglais au choix de l'auteur, traduits en anglais (0095-4527)
Fondation Wikimédia. 2010.
Voyez ce qu'est « Cellule (biologie) » dans d'autres dictionnaires :
LA BIOLOGIE- LA BIOLOGIE. Contenu : I. Histoire de la biologie.............. 424 Vitalisme et machinisme. L'émergence des sciences empiriques aux XVIe et XVIIIe siècles. L'émergence et le développement de la théorie évolutionniste. Développement de la physiologie au XIXe siècle. Développement de la science cellulaire. Résultats du 19ème siècle... Grande encyclopédie médicale
- (cellula, cytus), l'unité structurelle et fonctionnelle de base de tous les organismes vivants, un système vivant élémentaire. Peut exister en tant que département. organisme (bactéries, protozoaires, certaines algues et champignons) ou dans les tissus d'animaux multicellulaires,... ... Dictionnaire encyclopédique biologique
Les cellules des bactéries aérobies sporulées sont en forme de bâtonnet et, par rapport aux bactéries non sporulées, sont généralement plus grandes. Les formes végétatives de bactéries porteuses de spores ont un mouvement actif plus faible, bien qu'elles... ... Encyclopédie biologique
Ce terme a d'autres significations, voir Cellule (significations). Cellules sanguines humaines (HBC) ... Wikipédia
La cytologie (du grec κύτος formation en forme de bulle et λόγος mot, science) est une branche de la biologie qui étudie les cellules vivantes, leurs organites, leur structure, leur fonctionnement, les processus de reproduction cellulaire, le vieillissement et la mort. Les termes cellulaires sont également utilisés... Wikipédia
informations générales
La théorie cellulaire est une théorie fondamentale de la biologie, formulée au milieu 19ème siècle, qui a servi de base à la compréhension des lois du monde vivant et au développement doctrine évolutionniste. Matthias Schleiden Et Théodore Schwann formulé théorie cellulaire, basé sur de nombreuses études sur cage (1838 ). Rudolf Virchow plus tard ( 1858 ) l'a complété par la position la plus importante (chaque cellule provient d'une autre cellule).
Schleiden et Schwann, résumant les connaissances existantes sur la cellule, ont prouvé que la cellule est l'unité de base de tout corps. Cellules animaux, plantes Et bactéries ont une structure similaire. Plus tard, ces conclusions sont devenues la base pour prouver l'unité des organismes. T. Schwann et M. Schleiden ont introduit dans la science le concept fondamental de la cellule : il n'y a pas de vie en dehors des cellules. La théorie cellulaire a été complétée et éditée à chaque fois.
Dispositions de la théorie cellulaire de Schleiden-Schwann
Tous les animaux et plantes sont constitués de cellules.
Les plantes et les animaux grandissent et se développent grâce à l’émergence de nouvelles cellules.
Une cellule est la plus petite unité d’êtres vivants et un organisme entier est un ensemble de cellules.
Dispositions fondamentales de la théorie cellulaire moderne
Cellule- Il s'agit d'une unité élémentaire et fonctionnelle de la structure de tous les êtres vivants. (Sauf virus qui n'ont pas de structure cellulaire)
Cellule- un système unique, il comprend de nombreux éléments naturellement interconnectés, représentant une formation intégrale constituée d'unités fonctionnelles conjuguées - les organites.
Cellules de tous les organismes homologue.
Une cellule naît uniquement en divisant la cellule mère.
Un organisme multicellulaire est un système complexe de nombreuses cellules unies et intégrées dans des systèmes de tissus et d'organes connectés les uns aux autres.
Cellules d'organismes multicellulaires totipotent.
Une cellule ne peut provenir que d’une cellule précédente.
Dispositions supplémentaires de la théorie cellulaire
Pour rendre la théorie cellulaire plus conforme aux données de la biologie cellulaire moderne, la liste de ses dispositions est souvent complétée et élargie. Dans de nombreuses sources, ces dispositions supplémentaires diffèrent ; leur ensemble est assez arbitraire.
Cellules procaryote Et eucaryotes sont des systèmes de différents niveaux de complexité et ne sont pas complètement homologues les uns aux autres (voir ci-dessous).
La base de la division cellulaire et de la reproduction des organismes est la copie d'informations héréditaires - les molécules d'acide nucléique (« chaque molécule d'une molécule »). Les dispositions sur la continuité génétique s'appliquent non seulement aux cage dans son ensemble, mais aussi à certains de ses composants plus petits - à mitochondries, chloroplastes, gènes Et chromosomes.
Un organisme multicellulaire est un nouveau système, un ensemble complexe de nombreuses cellules, unies et intégrées dans un système de tissus et d'organes, reliés entre eux par des facteurs chimiques, humoraux et nerveux (régulation moléculaire).
Les cellules multicellulaires sont totipotentes, c'est-à-dire qu'elles ont le potentiel génétique de toutes les cellules d'un organisme donné, sont équivalentes en termes d'informations génétiques, mais diffèrent les unes des autres par l'expression (fonction) différente de divers gènes, ce qui conduit à leur morphologie et fonctionnalité diversité - à la différenciation.
17ème siècle
1665 - Physicien anglais R. Hooke dans son ouvrage « Micrographie », il décrit la structure du liège, sur des coupes minces dont il a trouvé des vides correctement localisés. Hooke a appelé ces vides « pores ou cellules ». Il connaissait la présence d'une structure similaire dans d'autres parties des plantes.
Années 1670 - Médecin et naturaliste italien M. Malpighi et naturaliste anglais N. a grandi ont décrit des « sacs ou vésicules » dans différents organes végétaux et ont montré la distribution répandue de la structure cellulaire dans les plantes. Les cellules ont été représentées dans ses dessins par un microscopiste néerlandais A. Levenguk. Il fut le premier à découvrir le monde des organismes unicellulaires - il décrivit les bactéries et les protistes (ciliés).
Les chercheurs du XVIIe siècle, qui ont montré la prédominance de la « structure cellulaire » des plantes, n'ont pas apprécié l'importance de la découverte de la cellule. Ils imaginaient les cellules comme des vides dans une masse continue de tissu végétal. Grew considérait les parois cellulaires comme des fibres, c'est pourquoi il a inventé le terme « tissu », par analogie avec le tissu textile. Les études sur la structure microscopique des organes animaux étaient aléatoires et n'apportaient aucune connaissance sur leur structure cellulaire.
XVIIIe siècle
Au XVIIIe siècle, les premières tentatives ont été faites pour comparer la microstructure des cellules végétales et animales. K.F. Wolf dans son ouvrage « La Théorie de la Génération » (1759), il tente de comparer le développement de la structure microscopique des plantes et des animaux. Selon Wolf, l'embryon des plantes et des animaux se développe à partir d'une substance sans structure dans laquelle les mouvements créent des canaux (vaisseaux) et des vides (cellules). Les données factuelles citées par Wolff ont été interprétées de manière erronée par lui et n'ont pas ajouté de nouvelles connaissances à ce que savaient les microscopistes du XVIIe siècle. Cependant, ses idées théoriques anticipaient largement les idées de la future théorie cellulaire.
19ème siècle
Dans le premier quart du XIXe siècle, les idées sur la structure cellulaire des plantes se sont considérablement approfondies, associées à des améliorations significatives dans la conception du microscope (en particulier la création lentilles achromatiques).
Lien et Moldnhower établissent la présence de parois indépendantes dans les cellules végétales. Il s'avère que la cellule est une certaine structure morphologiquement distincte. En 1831 G. Mol prouve que même des structures végétales apparemment non cellulaires, telles que des tubes aquifères, se développent à partir de cellules.
F. Meyen dans « Phytotomy » (1830) décrit des cellules végétales qui « sont soit solitaires, de sorte que chaque cellule représente un individu spécial, comme on le trouve dans les algues et les champignons, soit, formant des plantes plus hautement organisées, elles sont combinées en masses plus ou moins importantes. ". Meyen souligne l'indépendance du métabolisme de chaque cellule.
En 1831 Robert Brun décrit cœur et suggère qu'il s'agit d'un composant permanent de la cellule végétale.
École Purkinje
En 1801, Vigia a introduit le concept de tissu animal, mais il a isolé les tissus sur la base d'une dissection anatomique et n'a pas utilisé de microscope. Le développement d'idées sur la structure microscopique des tissus animaux est principalement associé aux recherches de Purkinje, qui a fondé son école à Breslau.
Purkinje et ses étudiants (il convient de souligner en particulier G. Valentin) ont révélé sous la forme première et la plus générale la structure microscopique des tissus et des organes des mammifères (y compris les humains). Purkinje et Valentin ont comparé des cellules végétales individuelles avec des structures tissulaires microscopiques individuelles d'animaux, que Purkinje appelait le plus souvent « grains » (pour certaines structures animales, son école utilisait le terme « cellule »).
En 1837, Purkinje donna une série de conférences à Prague. Il y rendait compte de ses observations sur la structure des glandes gastriques, du système nerveux, etc. Le tableau joint à son rapport donnait des images claires de certaines cellules de tissus animaux. Néanmoins, Purkinje n’a pas pu établir l’homologie des cellules végétales et des cellules animales :
premièrement, par grains, il entendait soit les cellules, soit les noyaux cellulaires ;
d’autre part, le terme « cellule » était alors compris littéralement comme « un espace délimité par des murs ».
Purkinje a mené la comparaison des cellules végétales et des « grains » animaux en termes d’analogie, et non d’homologie de ces structures (comprenant les termes « analogie » et « homologie » au sens moderne).
L'école de Müller et l'œuvre de Schwann
La deuxième école où l'on étudiait la structure microscopique des tissus animaux était le laboratoire Johannes Müllerà Berlin. Müller a étudié la structure microscopique de la corde dorsale (notocorde) ; son élève Henlé a publié une étude sur l'épithélium intestinal, dans laquelle il décrit ses différents types et leur structure cellulaire.
C'est ici qu'ont été menées les recherches classiques de Theodor Schwann, qui ont jeté les bases de la théorie cellulaire. Le travail de Schwann a été fortement influencé par l'école Purkinje et Henlé. Schwann a trouvé le principe correct pour comparer les cellules végétales et les structures microscopiques élémentaires des animaux. Schwann a pu établir l'homologie et prouver la correspondance dans la structure et la croissance des structures microscopiques élémentaires des plantes et des animaux.
L’importance du noyau dans une cellule de Schwann a été motivée par les recherches de Matthias Schleiden, qui a publié son ouvrage « Materials on Phytogenesis » en 1838. C’est pourquoi Schleiden est souvent appelé le co-auteur de la théorie cellulaire. L'idée de base de la théorie cellulaire - la correspondance des cellules végétales et des structures élémentaires des animaux - était étrangère à Schleiden. Il a formulé la théorie de la formation de nouvelles cellules à partir d'une substance sans structure, selon laquelle, d'abord, un nucléole se condense à partir de la plus petite granularité et autour de lui se forme un noyau, qui est le producteur de cellules (cytoblaste). Cependant, cette théorie reposait sur des faits incorrects.
En 1838, Schwann publie 3 rapports préliminaires, et en 1839 paraît son ouvrage classique « Études microscopiques sur la correspondance dans la structure et la croissance des animaux et des plantes », dont le titre même exprime l'idée principale de la théorie cellulaire :
Dans la première partie de l'ouvrage, il examine la structure de la notocorde et du cartilage, montrant que leurs structures élémentaires – les cellules – se développent de la même manière. Il prouve en outre que les structures microscopiques d'autres tissus et organes du corps animal sont également des cellules tout à fait comparables aux cellules du cartilage et de la notocorde.
La deuxième partie du livre compare les cellules végétales et les cellules animales et montre leur correspondance.
Dans la troisième partie, des positions théoriques sont développées et les principes de la théorie cellulaire sont formulés. Ce sont les recherches de Schwann qui ont formalisé la théorie cellulaire et prouvé (au niveau des connaissances de l'époque) l'unité de la structure élémentaire des animaux et des plantes. La principale erreur de Schwann était l'opinion qu'il a exprimée, à la suite de Schleiden, sur la possibilité de l'émergence de cellules à partir de matière non cellulaire sans structure.
La structure des animaux, comme de tous les autres organismes, repose sur la cellule. Il s'agit d'un système complexe dont les composants sont interconnectés par diverses réactions biochimiques. La structure exacte d’une cellule particulière dépend des fonctions qu’elle remplit dans le corps.
Les cellules des plantes, des animaux et des champignons (tous eucaryotes) ont un plan structurel général. Ils ont une membrane cellulaire, un noyau avec un nucléole, des mitochondries, des ribosomes, un réticulum endoplasmique et un certain nombre d'autres organites et autres structures. Cependant, malgré leurs similitudes, les cellules animales possèdent leurs propres caractéristiques qui les distinguent à la fois des cellules végétales et des champignons.
Les cellules animales sont couvertes uniquement membrane cellulaire. Ils n’ont ni paroi cellulaire cellulosique (comme les plantes), ni paroi cellulaire chitineuse (comme les champignons). La paroi cellulaire est rigide. Ainsi, d’une part, il fournit une sorte de squelette externe (support) à la cellule, mais, d’autre part, il ne permet pas aux cellules végétales et fongiques d’absorber des substances par capture (phagocytose et pinocytose). Ils les sucent. Les cellules animales sont capables de cette méthode de nutrition. La membrane cellulaire est élastique, ce qui permet de modifier dans une certaine mesure la forme de la cellule.
Les cellules animales sont généralement plus petites que les cellules végétales et fongiques.
Cytoplasme- Il s'agit du contenu liquide interne de la cellule. C'est visqueux car c'est une solution de substances. Le mouvement constant du cytoplasme assure le mouvement des substances et des composants cellulaires. Cela favorise la survenue de diverses réactions chimiques.
La place centrale dans la cellule animale est occupée par un grand cœur. Le noyau possède sa propre membrane (enveloppe nucléaire), séparant son contenu du contenu du cytoplasme. La membrane nucléaire possède des pores à travers lesquels s'effectue le transport de substances et de structures cellulaires. À l'intérieur du noyau se trouve le jus nucléaire (sa composition est quelque peu différente de celle du cytoplasme), nucléole Et chromosomes. Lorsqu’une cellule se divise, les chromosomes s’enroulent et peuvent être vus au microscope optique. Dans une cellule qui ne se divise pas, les chromosomes ressemblent à des fils. Ils sont en « état de marche ». A cette époque, ils synthétisent différents types d’ARN, qui assurent ensuite la synthèse des protéines. Les chromosomes stockent des informations génétiques. Il s'agit d'un code dont la mise en œuvre détermine l'activité vitale de la cellule ; il est également transmis aux cellules filles lors de la division de la cellule mère.
Les mitochondries, le réticulum endoplasmique (RE) et le complexe de Golgi possèdent également une enveloppe membranaire. DANS mitochondries La synthèse d'ATP (acide adénosine triphosphorique) se produit. Une grande quantité d'énergie est stockée dans ses connexions. Lorsque cette énergie est nécessaire à la vie de la cellule, l’ATP va progressivement se décomposer pour libérer de l’énergie. Sur PSE on retrouve souvent ribosomes, la synthèse des protéines se produit sur eux. Grâce aux canaux EPS, les protéines, les graisses et les glucides s'écoulent vers Complexe de Golgi, où ces substances s'accumulent puis sont libérées sous forme de gouttelettes entourées d'une membrane, selon les besoins.
Les ribosomes n'ont pas de membrane. Les ribosomes sont l’un des composants les plus anciens de la cellule, tout comme les bactéries. Contrairement aux eucaryotes, les cellules bactériennes ne possèdent pas de véritables structures membranaires.
Dans une cellule animale, il y a lysosomes, qui contiennent des substances qui décomposent la matière organique absorbée par la cellule.
Contrairement à une cellule végétale, une cellule animale ne possède pas de plastes, notamment de chloroplastes. En conséquence, la cellule animale n’est pas capable de se nourrir de manière autotrophe, mais se nourrit de manière hétérotrophe.
Dans une cellule animale, il existe des centrioles (centre cellulaire), qui assurent la formation du fuseau et la divergence des chromosomes lors de la division cellulaire. Une cellule végétale ne possède pas une telle structure cellulaire.




